
はじめに
東京大学大学院の講義「脳型情報処理機械論」の2回目だ。次回は「身体化された知性のためのフィジカル・リザーバー・コンピューティング」と言うテーマで東京大学AIセンター特任教授の中嶋浩平博士による講義だ。今回は次の4つの資料に基づく盛り沢山の講義だったので、正直消化不良気味だ。じっくりと復習をかねて頭の整理をした上で、何回かに分けて投稿したい。今回は特に視覚系の話を2回に分けて、その前半として投稿したい。今回も講義の内容をベースにネットで確認しながら理解したことをまとめるパターンである。なので、ここで記載したことの文責は筆者にあり、もし間違っていたり、不適切な内容があれば、筆者の責任である。逆に、内容が分かりやすいとすればそれは國吉教授の講義のおかげである点を強調しておきたい。
・視覚系の情報の流れ ⇨ 今回は視覚系を2回に分け、そのうちの前半。後半は別途。
・脳の運動制御系 ⇨別途。
・脳の仕組み、ロボット ⇨別途
(英語タイトル:Development of Embodied Brain, Behavior and Cognition)
視覚系の情報の流れ
視覚経路と視覚野
右目の網膜で検出した信号は左脳へ、左目の網膜で検出した信号は右脳にて処理される。また、視覚には、形態視系と運動視系がある。前者は、ParvoCellular(P-Channnel)で下の図(右)の赤い線が4本ある。後者は早い動きに追随するMagnoCellular(M-Channel)で下の図(右)の青い線で2本ある。網膜で検出した信号はLGNを経由して一次視覚野(V1)を通って、形態視系はDorsal Pathwaryに向かい、運動視系はV1からVentral Pathwayに向かう。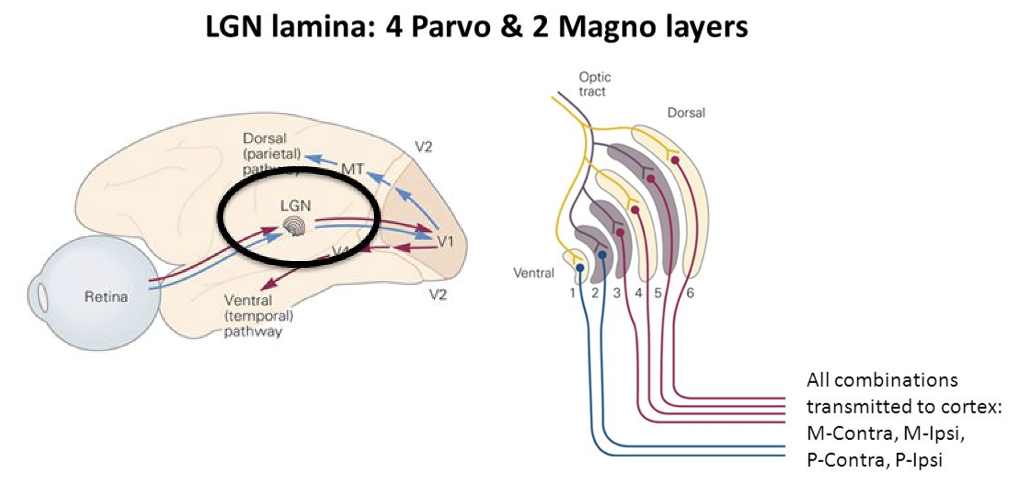
(出典:Slideplayer)
一次視覚野
前述の通り、視覚系には形態視系と運動視系の機能分化がある。前者は、一次視覚野(V1)を起点として頭頂葉に向かう背側経路であり、空間知覚や動きの情報処理に関わる。一方、後者はV1から下頭側皮質に向かう腹側経路であり、物体認知に関わる。腹側経路はV1からV2野、V4野を経て、下頭側皮質に向かう経路であり、色情報はもっぱらこの経路で処理されている。
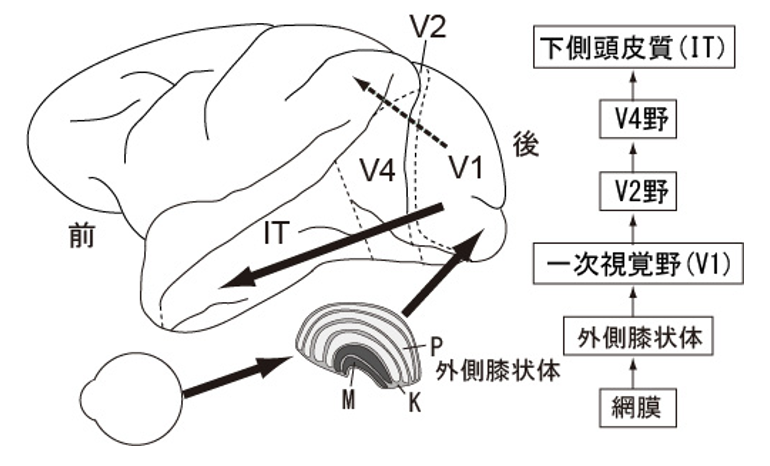
(出典:色選択性細胞)
視覚前野と下側頭皮質
V1で処理された色情報は、大脳視覚野の腹側経路を構成するV2野、V4野を経て、下頭側皮質に伝えられる。V2野ではチトクロムオキシダーゼ染色で濃く染まる細い縞の領域(thin stripe)内に多く見られる。V4野と下頭側皮質では、色刺激に強く応答する数ミリ程度のサイズの小領域が複数存在する。ヒトは腹側後頭葉皮質の紡錘状回付近の損傷により色知覚に重篤な障害が生じることがあり、大脳性色覚異常とよばれる。
(出典:bsd)
ゼロ視差フィルター
下の図(左)は赤い点と青い点と緑の点を両眼で見ている様子だ。左目では位置はほぼ同じだが奥行きが異なる。一方、右目では赤い点は中心点に近いが、青、緑と徐々に遠のく。この左右の網膜を見かけ上並行にして、上と下で対比できる容易に描いたものが下の図(右)だ。注視点と同じ奥行きにある青い点のの左右の像は、中心窩を基準とした網膜座標上の同じ位置に投影され、その両眼視差はゼロとなる。一方、注視点と異なる奥行き面上にある刺激である赤い点と緑の点は、左右網膜の異なる位置に投影され、ゼロ以外の両眼視差をもつ。手前にある緑の点と、奥にある赤い点の両眼視差の方向は逆になる。
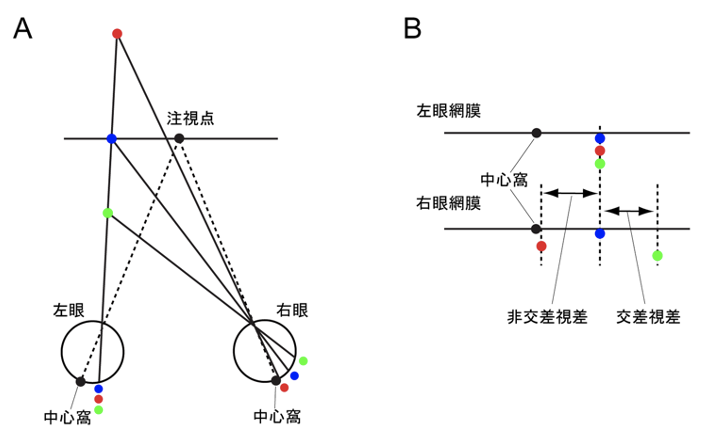
(出典:両眼視差)
方向選択性
一次視覚野(V1)で見られる色選択性の特性は、色空間のさまざまな方向にチューニングを持つニューロンが見られることである。色空間の方向によって色相が変化するので、V1の細胞はさまざまな色相に選択性を持つ。大脳腹側視覚経路の各領野ではさまざまな色相に選択的に応答するニューロンが共通して見られるが、そのような反応特性の形成はV1の段階で始まる。大脳皮質視覚野がニューロン線分に反応するニューロンを3つに分類できる。単純型細胞は光がついたときに反応するONの受容野と、光が消えたときに反応するOFFの受容野、そして、受容野の全ての場所でON反応とOFF反応を示す複雑型細胞だ。超複雑型細胞は、複雑型細胞の性質に加えて、線分が長くなると反応しなくなる性質を持つ。これら3種類のニューロンは、ニューロンごとに好みの傾きを持っている。下の図に示すように、ニューロンは、45度の傾きの長方形が左下から右上に動いたときに最も強く反応した。
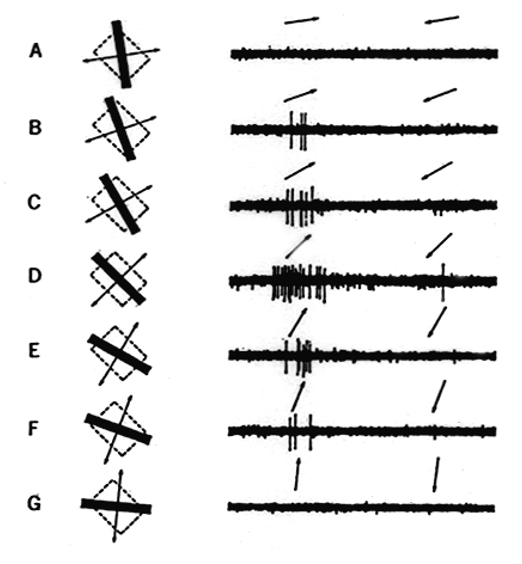
(出典:脳の世界メニュー)
V1情報表現
一次視覚野(V1)は視覚における空間情報のマップを持ち、ヒトの鳥距溝上壁は視野の下半分の視覚情報に強く応答し、下壁は視野の上半分に応答する。視野と V1 の位置の対応は非常に正確である。中心窩を持つ動物やヒトでは、V1 の大きな割合を占める領域が、視野の中心にある小さい領域をマップし、この現象は皮質拡大(cortical magnification)という。V1 ニューロンの応答選択性は時間によって大きく異なり、40ms前後の短時間で、 V1 ニューロンは刺激の小さな組に対して強くチューニングされる。V1 と一般的な一次感覚野のニューロンは似たチューニング特性を持つもので集まり、皮質コラムと呼ばれる。現在、V1 ニューロンの初期の応答は選択的な時空間フィルター(spatiotemporal filters)で構成されていると考えられている。時間的に100ms前後と遅いV1 のニューロンはシーンのより大局的な構造に応答すると考えれている。
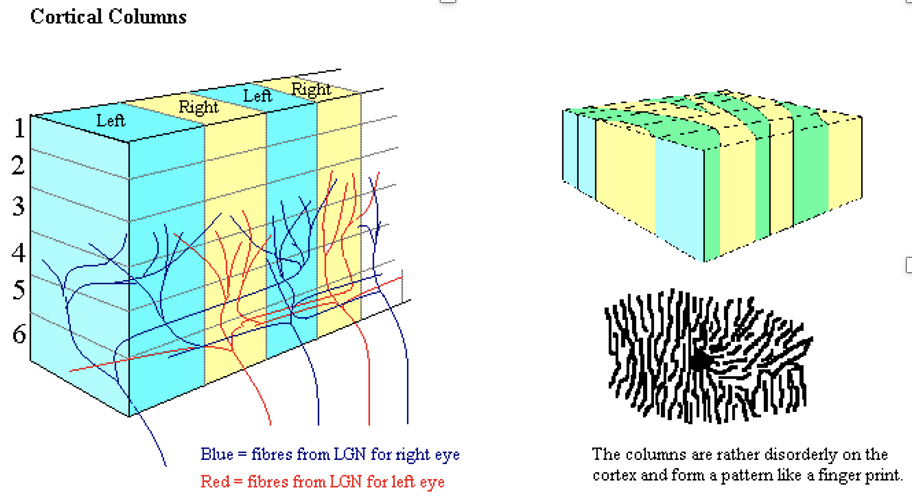
(出典:視覚野)
眼優位性
下の図は、猫の視覚伝導路を示す模式図だ。視野の半分(実線部分)の情報は両眼で捉えられた後、一側の外側膝状体の異なる層に伝達される。外側膝状体ニューロンはV1のⅣ層に投射する。2つの眼で捉えた視覚情報は脳で統合して、1つの視覚イメージが出来上がる。それぞれの眼球を出た視神経は視交叉で融合し、すぐ再び左右に分離して視索となる。視神経軸索の一部は交差して対側の脳に向かい、残りは同側に向かう。左右の網膜の、視野上で対応する部分に由来する情報は、脳の同じ部位に収束する。このような部分交差のアイデアはアイザック・ニュートンが最初に提唱したとされる。両眼からの入力が収束することで両眼に反応するニューロンが生まれるが、それは大脳皮質一次視覚野で初めて観察される。個々のニューロンがどちらの眼により強く反応するかを眼優位性という。このように眼優位性は視覚野ニューロンの反応特徴であって、いわゆる「利き目(dominant eye)」のことではない。
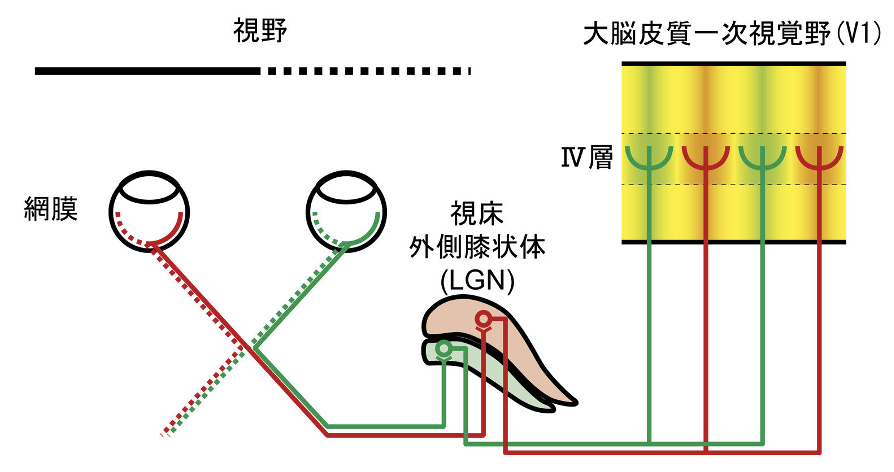
(出典:眼優位性)
コラム構造
網膜と第一次視覚野(V1)の間には連続的な 1 対 1 対応が存在し(レティノトピー:retinotopy)、鼓膜の周波数選択特性と第一次聴覚野(A1)との間にも対応関係が見られる(トノトピー:tonotopy)。 同様に体表面の感覚と体制感覚野の間にも対応関係が見られる(ソマトピー:somatopy)。つまり、感覚器官と第一次感覚野との間の神経結合は 類似した刺激に対し、皮質上の同じような位置に対応する受容野を持つ。 このような 2 つの神経場間の連続的な結合関係のことをトポグラフィックマッピング(topographic mapping)と言う。視覚野のトポグラフィックマッピングについては、任意の視覚位置に対して眼優位性(ocular dominancy)や方位選択性(orientation selectivity)、色などの情報が処理されるように規則正しく配列されている。これをハイパーコラム(hypercolumn)構造という。
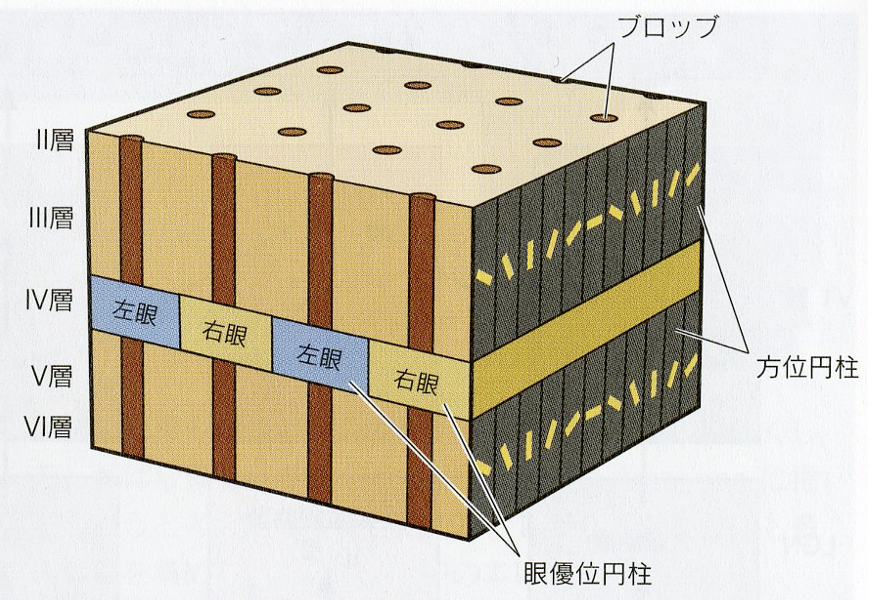
(出典:東京女子大学)
ピンウィール(Pin Wheel:風ぐるま)
下の図は、ネコの視覚野のピンウィール内にある数千個のニューロンの方位同調曲線(pinwheels in the visual cortex)だ。ピンウィールは、好ましい刺激の向きが異なる多数の細胞が、技術的には特異点と呼ばれる点に集まっている領域である。ピンウィールは、刺激の方向を異にする細胞が多数集まった領域であり、専門的には特異点と呼ばれている。ピンウィールを可視化するために用いられる光学イメージング技術は,多数のニューロンからの信号を平均化するものである。ピンウィールセンターでは、細胞が単に方向性に反応しなくなったり、選択されなくなったりする可能性もあるが、それを否定する研究もある。
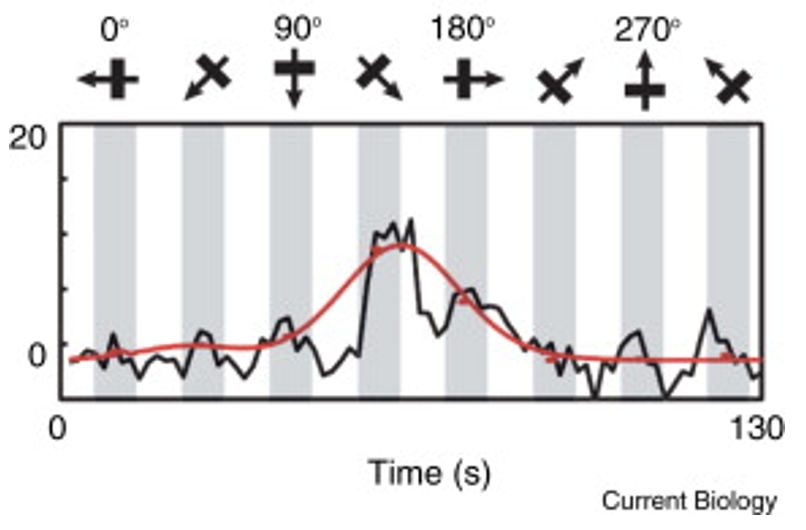
(出典:Current Biology)
まとめ
國吉教授の「脳型情報処理機械論」の第1回講義については、脳の話とニューロンの話の2つに分けてすでに投稿した。今回は、視覚系の情報の流れを中心にまとめてみた。後頭部にある一次視覚野(V1)に情報が集まり、そこから静態情報を大脳の方に、動態情報は下頭側皮質に向かう。色の情報もこの経路で使うようだ。男性は動くものが好きだ。女性のポニーテールがゆらゆらと揺れると男性は本能的に気になるし、可愛いと感じるものだ。また、大脳皮質の表面は六層構造をしており、眼優位性を示すニューロンが表面と垂直に並び、宝庫コラムを形成している。これはまるで量子コンピュータではないか。また、ピンウィール(Pinwheel)は眼優位性コラムの中央部に配列されるが、時計回りのピンウィールと反時計回りのピンウィールが存在するのも、量子スピンのようだと思った。
以上
最後まで読んで頂きありがとうございました。


