
- 1 はじめに
- 2 4つの論点
- 3 論点1:ニューロンはどのように生成されたか?
- 3.1 神経細胞(Neurons)
- 3.2 細胞分化(cellular differentiation)
- 3.3 神経前駆細胞(neural progenitor cell)
- 3.4 グリア前駆細胞(Glial restricted progenitor cells:GRP)
- 3.5 増血幹細胞の老化(Hematopoietic Aging)
- 3.6 幹細胞生物学の力学的システムビュー(A Dynamical-systems view of Stem Cell Biology)
- 3.7 脱分極(Depolarization)
- 3.8 スパイキングニューラルネットワーク(Spiking Neural Network:SNN)
- 4 論点2:機能モジュールはどのように生成されたのか?
- 5 まとめ
はじめに
前回は、脳型情報処理機械論の第4回講義のその1として、複雑系カオス脳理論を論じる上での前提となる基本的な知識などをまとめた。今回は、津田教授が提示した4つの論点のうち最初の2つについての考察を理解した範疇でまとめてみた。なお、今回の投稿も、あくまで津田教授の講義を聞いた中で興味深いと感じたことをネットで確認しながらまとめたものであり、講義メモではない点はご承知おきください。
その1:4つの論点と前提となる知識 (前回の投稿)
その2:4つの論点(前半)のレビュー(⇨ 今回)
その3:4つの論点(後半)のレビュー(次回)
4つの論点
論点1:ニューロンはどのように生成されたか?
論点2:機能モジュールはどのように生成されたのか?
論点3:神経細胞はどのようにして機能特異性を獲得したのか?
論点4:海馬・記憶システムの進化
論点1:ニューロンはどのように生成されたか?
ニューロンについては、この最初の講義を受けた時の投稿でまとめたが、人間の脳には約1000億個のニューロン(神経細胞:Neuron)が存在する。論点1はそのニューロンはどのように生成されたのか?それを順番に紐解いていきたい。
神経細胞(Neurons)
神経細胞とは、神経系に存在する細胞であり、体内の他の細胞を刺激してコミュニケーションをとるための特殊な細胞だ。神経細胞は興奮性で、電気的な刺激を受けて機能する。活動電位と呼ばれる電気的メッセージを通じて、神経細胞は標的とする細胞に作用を引き起こす。ニューロンには大きく分けて、感覚ニューロン、中継ニューロン、運動ニューロンの3種類がある。
感覚ニューロン(Sensory Neurons)
目、耳、舌、皮膚などの受容体に存在し、神経インパルスを脊髄や脳に伝える。これらの神経インパルスが脳に到達すると、視覚、聴覚、味覚、触覚などの感覚に変換される。しかし、すべての感覚ニューロンが脳に到達するわけではなく、一部のニューロンは脊髄に伝達され、迅速な反射行動を可能にする。
リレーニューロン(Relay Neurons)
感覚入力と運動出力の間にあるのがリレーニューロンだ。リレーニューロンは脳と脊髄に存在し、感覚ニューロンと運動ニューロンのコミュニケーションを司る。第2回の講義で投稿したように、形態視系の情報は前頭葉に向かうが、運動視系の情報は下頭側皮質から脊髄に向かい、反射的な動きを可能としている。
運動ニューロン(Motor Neurons)
運動ニューロンは、中枢神経系(CNS:central nervous system)に存在し、筋肉の動きを制御する。運動ニューロンが刺激を受けると、神経伝達物質が放出され、それが筋肉上の受容体に結合して反応を起こし、運動につながる。
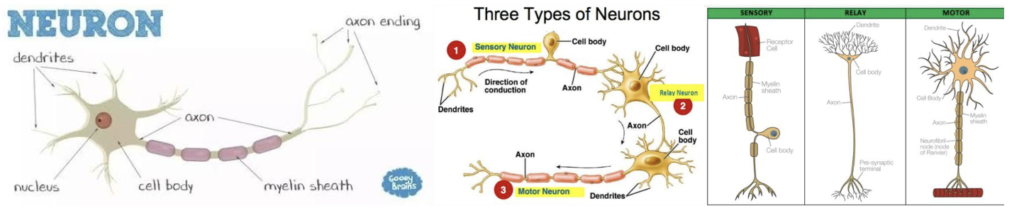
(出典:quota)
細胞分化(cellular differentiation)
細胞分化(cellular differentiation)とは,細胞がある種類の細胞から別の種類の細胞へと変化する過程である。通常,細胞はより特殊な種類の細胞へと変化する。分化は、多細胞生物が単純な接合体から組織と細胞タイプの複雑なシステムへと変化する過程で何度も起こる。分化は大人になっても続き、組織の修復や正常な細胞のターンオーバーの際に、成体幹細胞が分裂して完全に分化した娘細胞を作り出す。一部の分化は、抗原の暴露に反応して起こる。分化は、細胞の大きさ、形状、膜電位、代謝活動、シグナルに対する反応性を劇的に変化させる。脊椎動物の神経系、筋骨格、表皮、腸などの一部の組織では、終末分化と呼ばれる特殊なタイプの分化が重要な意味を持つ。末端分化では、以前は細胞分裂が可能であった前駆細胞が細胞周期から永久に外れ、細胞周期装置を解体し、細胞の最終的な機能に特徴的な一連の遺伝子を発現することが多い。最終分化後も、細胞の能力や機能がさらに変化する場合には、分化が継続することがある。分裂した細胞の中には、他の種類の細胞に分化する能力である細胞の力価が複数のレベルで存在する。力価が高いほど、分化できる細胞の種類が多いことを意味する。胎盤組織を含むすべての種類の細胞に分化できる細胞を「全能性」という。単能性細胞は、1つの細胞型にしか分化しないが、自己再生が可能だ。
神経前駆細胞(neural progenitor cell)
複雑な大脳皮質は哺乳類の脳の最大の特徴とされるが、これは複数の神経前駆細胞が多様な神経細胞を産生したためだ。発生期における大脳皮質の神経上皮に生じた神経幹細胞が分裂して数を増やし、やがて未分化型前駆細胞(放射状グリア細胞あるいはapical progenitor)、中間型前駆細胞(basal progenitorあるいはintermediate progenitor)、およびoRG前駆細胞(OSVZ (outer subventricular zone) radial glia-like cell)などの前駆細胞が出現する。これらの細胞は異なる分裂・分化能力を有し、固有の役割を担う結果として、組織における複雑な細胞構築が可能だ。
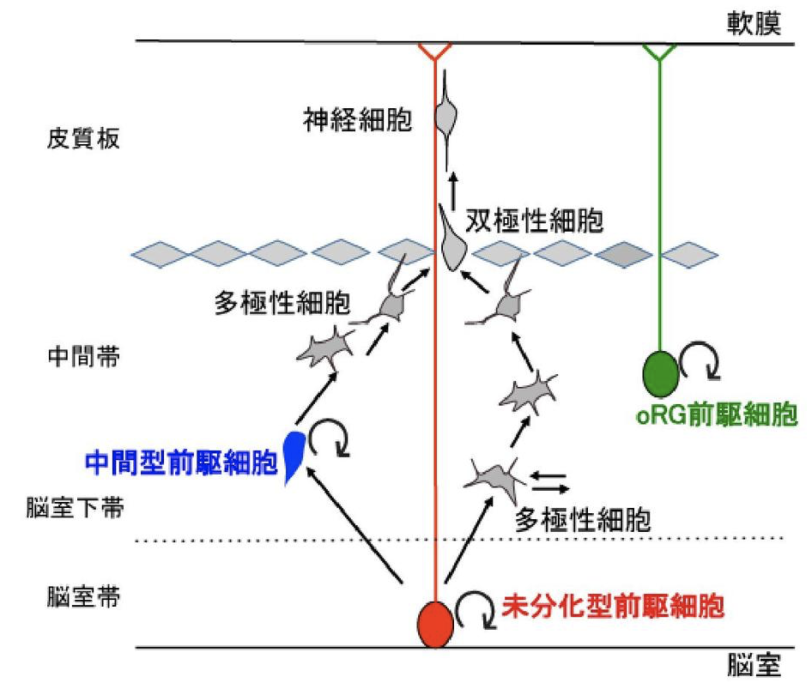
(出典:神経前駆細胞)
グリア前駆細胞(Glial restricted progenitor cells:GRP)
グリア制限前駆体細胞(GRP)が最も初期のグリア前駆細胞ならすべてのグリアの祖先であるかもしれない。NEP細胞がさまざまなグリア前駆細胞を生み出し、異なる種類の前駆細胞を生み出しているのかもしれない。GRP細胞がオリゴデンドロサイトや1型および2型のアストロサイトの表現型を持つ細胞を作り出す能力を持つことと一致する。さらなる検証が必要だが、GRP細胞が最も初期のGRP細胞を表しているようだ。これらの細胞は、同じく最近E13.5脊髄から分離されたニューロン制限前駆体細胞の補完的な前駆体細胞である。造血幹細胞から骨髄系とリンパ系が早期に分離することが提唱されているのと同様に、NEP細胞由来のGRPと神経細胞制限前駆体細胞は、CNSの正統的な系統制限された芽細胞である可能性があると考えられる。
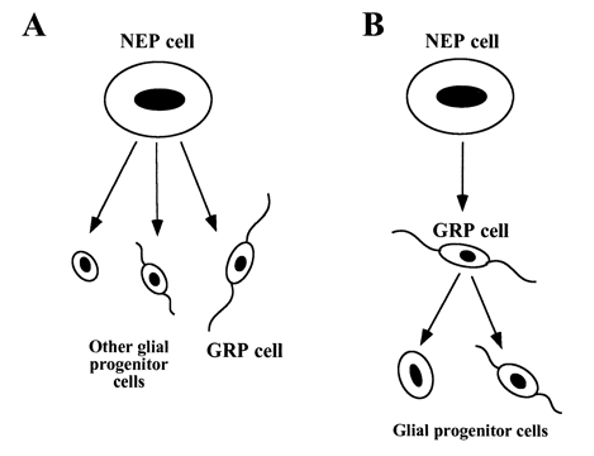
(出典:pnas.org)
増血幹細胞の老化(Hematopoietic Aging)
平均寿命が延び続けている。加齢に伴う病気は、個人レベルの問題であると同時に、社会経済レベルの問題だ。血液細胞の形成過程に関する研究から、加齢に伴ういくつかの欠点が明らかになった。例えば、適応免疫反応の能力低下や、血液細胞の組成が骨髄系細胞に偏ることなどが挙げられる。後者は癌を含む骨髄系疾患の発生率が劇的に増加する懸念がある。逆に、小児の急性白血病の多くはリンパ系細胞に起因する。最近のデータでは、幹細胞の加齢表現型は少なくともある程度可逆的である。つまり、高齢者の幹細胞の老化現象を若返らせたり、老化現象を弱める治療の効果が期待できる可能性を示唆されている。人類はどこまで長生きするのだろう。
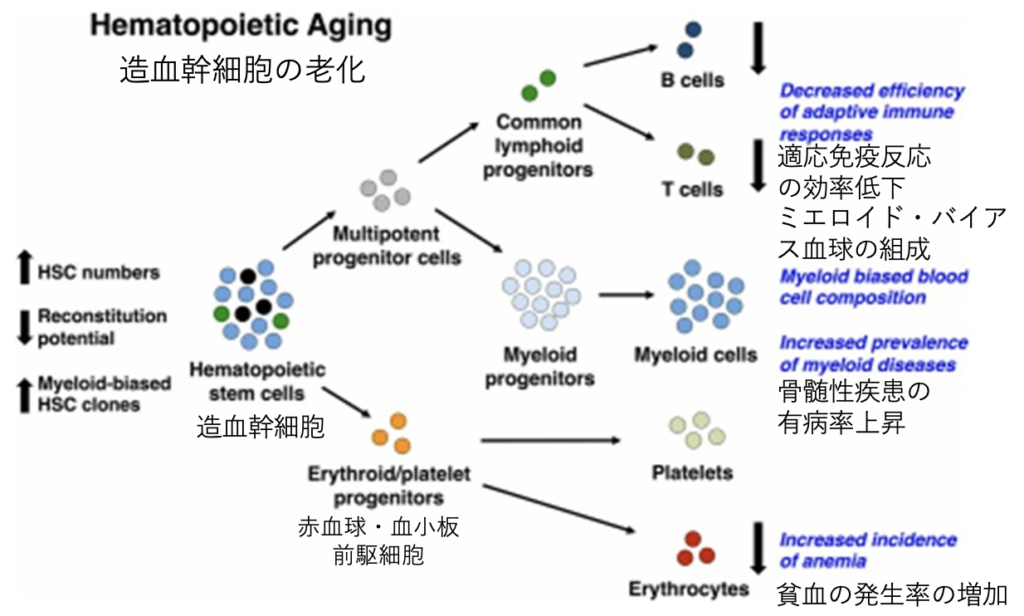
(出典:stem cells journals)
幹細胞生物学の力学的システムビュー(A Dynamical-systems view of Stem Cell Biology)
発生過程において、細胞は一方向の分化を経て、なりうる細胞の種類を徐々に減らしていく。しかし、幹細胞は、増殖と分化の両方の可能性を持っている。そのため、幹細胞は他の細胞種と異なり、安定した増殖と分化を行うことができるが、この特徴を理解することが非常に重要な課題となる。幹細胞は、それぞれが特有の性質である遺伝子発現レベルの変動や振動を有していることが判明している。
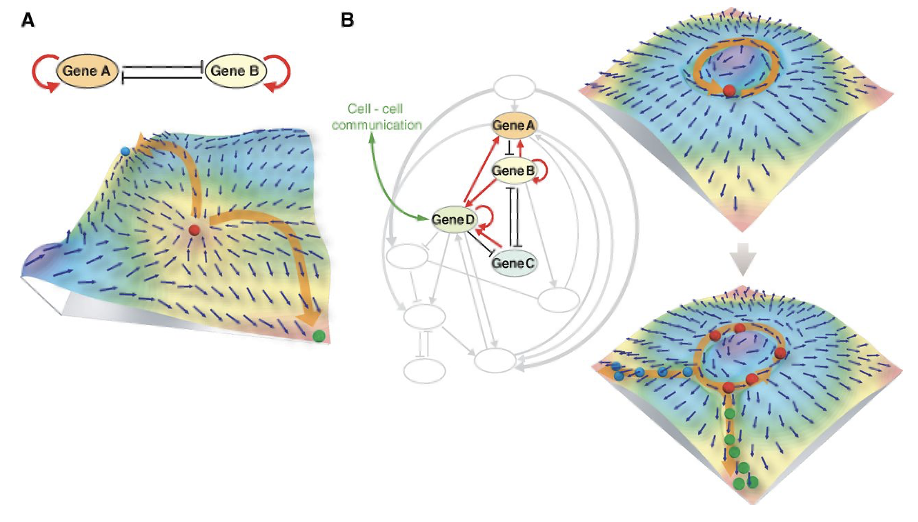
(出典:Science)
脱分極(Depolarization)
脱分極(Depolarization)とは、細胞内の変化のことで、その間、細胞は電荷分布の変化を受け、細胞内の負電荷が外部に比べて少なくなる。脱分極は、多くの細胞の機能、細胞間のコミュニケーション、および生物の全体的な生理機能に不可欠である。高等生物のほとんどの細胞は、外部に対して負の電荷を帯びた内部環境を維持している。この電荷の差は、細胞の膜電位と呼ばれる。脱分極の過程では、細胞の負の内部電荷が一時的に正になる。この膜電位のマイナスからプラスへの変化は、活動電位をはじめとするいくつかの過程で起こる。活動電位では、脱分極が非常に大きいため、細胞膜の電位差が一時的に反転し、細胞内部が正に帯電する。電荷の変化は、典型的にはナトリウムイオンの細胞内への流入によって起こるが、あらゆる種類の陽イオンの流入またはあらゆる種類の陰イオンの流出によって媒介されうる。脱分極の反対は過分極と呼ばれる。
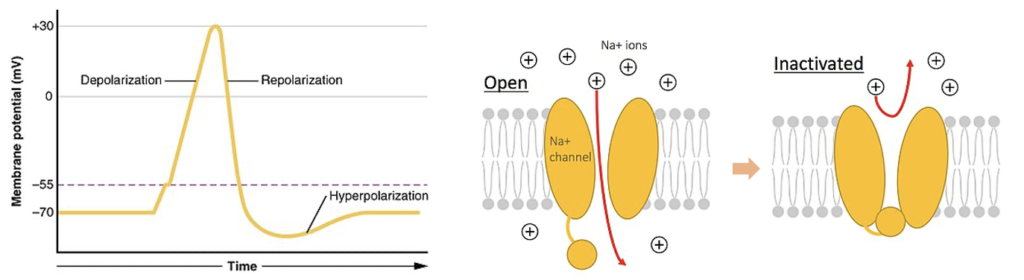
(出典:Depolarization)
スパイキングニューラルネットワーク(Spiking Neural Network:SNN)
スパイキングニューラルネットワーク(SNN)は、自然界のニューラルネットワークにより近い人工的なニューラルネットワークである。SNNは、ニューロンやシナプスの状態に加えて、時間の概念を動作モデルに組み込んでいる。SNNのニューロンは、一般的な多層パーセプトロンネットワークのように伝搬サイクルごとに情報を伝達するのではなく、ニューロンの膜の電荷に関連する固有の性質である膜電位が閾値と呼ばれる特定の値に達したときにのみ情報を伝達する。膜電位がしきい値に達すると、ニューロンが発火して信号が発生し、他のニューロンに伝わり、ニューロンはこの信号に応じて膜電位を増減させることになる。閾値を超えた瞬間に発火するニューロンモデルは、スパイキングニューロンモデルとも呼ばれる。最も代表的なスパイキングニューロンモデルは、リーキー・インテグレート・アンド・ファイア・モデルだ。積分発火モデルでは、通常、瞬間的な活性化レベルがニューロンの状態とみなされ、入力されるスパイクによってこの値が高くなったり低くなったりして、最終的に状態が減衰するか、発火閾値に達した場合にニューロンが発火するまで続く。発射されたスパイク列を実数として解釈するために、スパイクの頻度、刺激後の最初のスパイクまでの時間、スパイク間の間隔のいずれかに基づいて、さまざまなデコーディング方法が存在する。

(出典:Spiking Neural Network)
論点2:機能モジュールはどのように生成されたのか?
ニューロンがどのように生成したのかは論点1でまとめたが、それらのニューロンはどのように組織化されて機能を持つようになったのだろう。
ニューロンシステムのモジュール構成
人間の脳は、スケールフリーの高結合モジュールが、弱いリンクで疎結合されてスモールワールドネットワークを形成する複雑な構造をしているという。下の図は、M=14個のモジュールを異なる色で示したネットワークで、ニューロンの数はNm=20から100だ。モジュール内の結合は、モジュール間の結合よりも平均強度が大きい。実験データで観察されたスケーリング行動を非モジュール型ネットワーク上で再現することに成功した。このモデルのニューロン活動を、ヒトの脳で測定された主な統計的特徴を実装したモジュール型ネットワーク上で分析した結果、モジュール型ネットワークでは、シナプス結合の強さやモジュールの大きさや数にかかわらず、活動は完全にはスケールフリーにならないことがわかったという。

(出典:nature)
人工ニューラルネットワーク(Artificial Neural Network:ANN)
人工ニューラルネットワーク(ANN)は、動物の脳を構成する生物学的なニューラルネットワークにヒントを得て開発された計算システムである。下の図(左)は脳のニューロンを単純化して考え、相互に接続されたノード群であり、円形のノードが人工ニューロンを表し,矢印はある人工ニューロンの出力から別の人工ニューロンの入力への接続を表している。下の図(右)はニューロンと有髄の軸索、樹状突起の入力から軸索の末端の出力までの信号の流れである。ANNは、人工ニューロンと呼ばれる接続されたユニットまたはノードの集合体に基づいており、生物の脳のニューロンを緩やかにモデル化する。各接続は、生物学的脳のシナプスのように、他のニューロンに信号を伝達する。人工ニューロンは信号を受け取り、それを処理して、接続されたニューロンに信号を送る。接続部の信号は実数であり、各ニューロンの出力は、入力の合計の非線形関数によって計算される。接続部分はエッジと呼ばれ、ニューロンとエッジには学習の進行に応じて調整されるウェイトがある。ウェイトは、接続部の信号の強さを増減させる。ニューロンには閾値があり、信号の総和がその閾値を超えた場合にのみ信号が送られる。一般的に、ニューロンは層別に集約される。異なる層では、入力に対して異なる変換を行う。信号は、最初の層から最後の層まで、複数の層を経由して送られる。
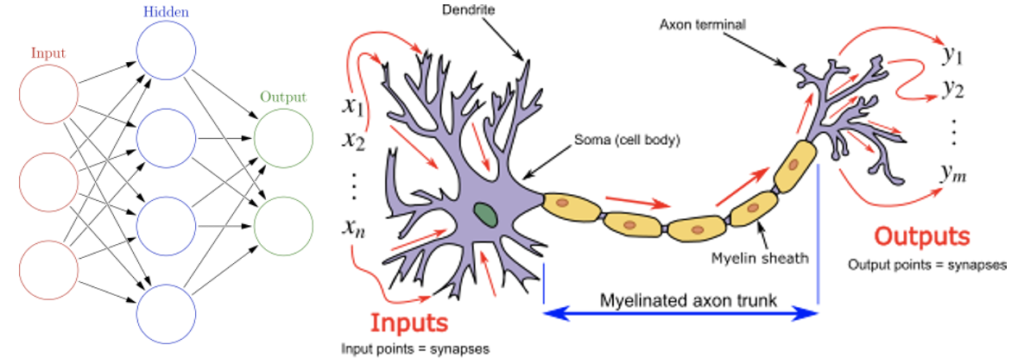
(出典:artificial neural network)
ホモジニアスなネットワークとヘテロジニアスなネットワーク(Homogeneous Network and Heterogeneous Network)
ホモジニアスなネットワークとは、ネットワーク内のすべてのノードが同じ機能を持つネットワークのことだ。つまり、全てのノードに個性がなく、同質だ。一方、ヘテロジニアス(不均質な)ネットワークとは、機能と効用の両方で分類された2つ以上のクラスのノードが存在するネットワークのことだ。組織として強いのはヘテロジニアスなネットワークだろう。それぞれのノードが個性を持ち、それぞれの強みを活かすようなら最高だ。
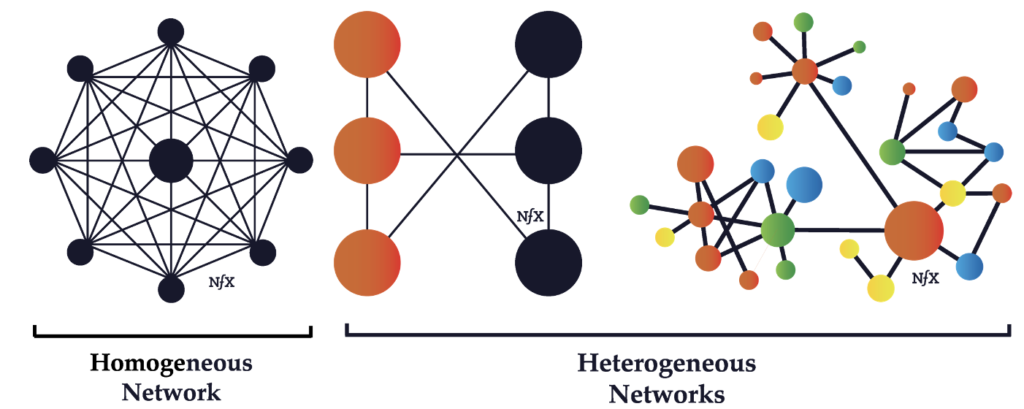
(出典:network effets bible)
カオスな遍歴(Chaotic itinerancy)
カオスな遍歴は、2013年に津田教授がコンピュータサイエンスに公開した内容だ。カオスな遍歴は、高次元力学系における普遍的な力学であり、低次元の秩序状態から高次元のカオスを経て、様々な状態を遍歴することを示すと言う。津田教授の論文では、カオス遍歴の発見、基本的特徴、特徴づけ、例示、意義などについて概説されている。
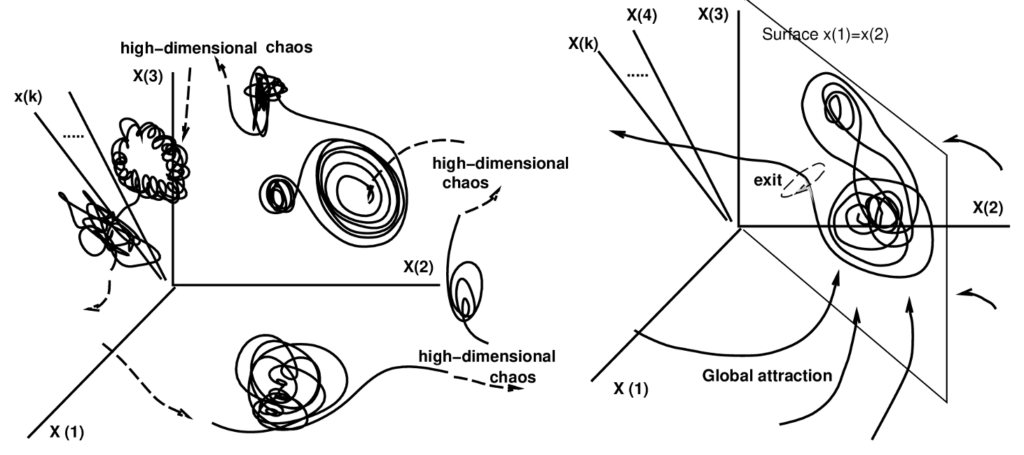
(出典:Semanticscholar)
まとめ
講義を聞いていても難解だったが、ネットや文献で色々調べて整理したつもりだけど、まだ難解だ。なぜ難解なのかというと、生物学的な観点と情報処理的な観点が交わるためだ。単に人間の脳の構造を分析するだけではなく、その意味を理解して、コンピュータネットワークに応用したいという狙いがある。それが問題を複雑にしているような気がする。かつて、鳥のように大空を飛びたいと多くの人がチャレンジして、その多くは失敗したが、最終的に成功したのは飛行機であり、ドローンである。人間には人間の生物としての強みと制約があり、コンピュータにはコンピュータの強みと制約がある。人間の脳の仕組みの優れた点とコンピュータの優れた点を高次元で組み合わせるような巧みさが求められているような気がする。
以上
最後まで読んで頂きありがとうございました。
拝
(参考) 4つの論点の後半2つは次回の投稿予定。
論点3:神経細胞はどのようにして機能特異性を獲得したのか?
論点4:海馬・記憶システムの進化
今後の検討課題


